
Guest Author: Jack Cooper, MSc
UoB Graduate / PhD Student, University of Swansea
As we’ve seen so far, there have been changes and updates to the scientific consensus of Megalodon’s taxonomic assignment and maximum body size. However, its favourite snack is something that is universally agreed upon in the scientific community. In another rarity, it’s something that’s usually portrayed with pretty good accuracy in fiction. Allow me to introduce a shark that ate whales – actual whales!
Many of today’s large macropredatory sharks are known for eating marine mammals, especially our old friend the great white shark. Some of the most iconic photographs of this shark include it hurling itself out of the water with its latest victim between its jaws (Fig. 1). However, juvenile white sharks don’t eat seals right from the get-go. Instead, they start off eating fish and smaller sharks [1]. If we look again at the Gatun formation in Panama, we find plenty of fossil sharks, rays and otoliths (structures from the inner ears of fish), as well a plethora of invertebrates like molluscs [2,3].
Something we do not find, however, is fossil marine mammals. This seems to indicate that Megalodon young were also eating fish and smaller sharks rather than immediately going after marine mammals. This is something called an ontogenetic shift in diet, meaning that the shark changes its preferred food source as it grows. By eating larger food, and probably lots of it, as it reached its adult size, Megalodon would’ve been able to maintain the metabolic demands of being so large. It could’ve done this by eating not only whales, but just about anything it came across.

Given how much food Megalodon would’ve needed to eat to justify its enormous size, it was almost certainly an opportunistic feeder. The fossil record is our best guide here. This is due to the presence of trace fossils, which are geological records of biological activity. This can range from dinosaur footprints to scratches on bones indicating that something bit the animal that bone belonged to. And Megalodon left a few of these trace fossils behind, providing key evidence of its kills.
Several fossils have been found belonging to a variety of cetaceans with notable bite marks on them. One well-cited example comes from the Yorktown formation in Maryland, where several whale bones had bite marks that had clearly been made by large serrated teeth [4]. In fact, Megalodon teeth are sometimes found directly associated with these bones [5].
Sharks lose and replace teeth thousands of times throughout their lives, and thus sharks frequently lose teeth when making a violent kill. This is a key reason why shark teeth are such common fossils. The presence of the shark’s teeth right by a scratched whale bone does indicate that feeding was going on. But, something that palaeontologists should always consider when these fossils are found is: was this actually predation? Or was it perhaps scavenging?
Scavenging is a common feeding behaviour in modern macropredatory sharks such as great whites (Fig. 2) and tiger sharks [6,7]. When these sharks encounter dead whales floating on the surface, it’s an all-you-can-eat buffet. Critically, scavenging requires no energy expenditure, something that can be wasted on a failed predation attempt. Given that Megalodon needed to be able to maintain its enormous size, it wouldn’t make much sense to pass on a free meal if it encountered a dead whale. Thus, by proxy of how large modern sharks feed, most of us would be willing to bet that Megalodon probably did scavenge when given a chance.

This said, there is one particularly remarkable fossil that implicates Megalodon in a predation attempt. And, crucially, we know this primarily because its attempt failed. A paper published in 2010 [8] described a partial rib Pliocene fossil belonging to a cetacean from the Yorktown formation in North Carolina (yep, two Yorktown formations). This fossil also showed damage resembling bite marks, but notably there appeared to be a woven bone covering over these marks. The authors interpreted this as a sign of an infection that had occurred while the animal had been recovering from bone trauma caused by those bite marks. These marks matched up well with a serrated bite, suggesting that the attacker had serrated teeth – just like Megalodon. The study therefore suggested that a Megalodon, or another large shark, had attacked the animal the fossil had come from, but the victim had survived the attempt only to die of an infection from its injury six weeks later [8].
Even more recently, the first ever fossil bite marks found in the southern hemisphere attributed to Megalodon were found in Peru’s Pisco formation [9]. Perhaps it was meant to be, as this is the same site where Gordon Hubbell found his exceptional Carcharodon fossil (discussed in Q1). The bitten material found here included skull and rib remains, suggesting that the shark had bitten its target from the side. Most significantly, the remains were identified to an individual species for the first time – that species being Piscobalaena nana, a small baleen whale. Like in the other trace fossils, the bite marks matched up well to large serrated teeth – a good case for accusing Megalodon of the attack (Fig. 3). It therefore appeared that Megalodon was also capable of targeting smaller whales (be it predation or scavenging), very much in line with suggestions that it likely also targeted dolphins, sea turtles and other smaller prey [9].

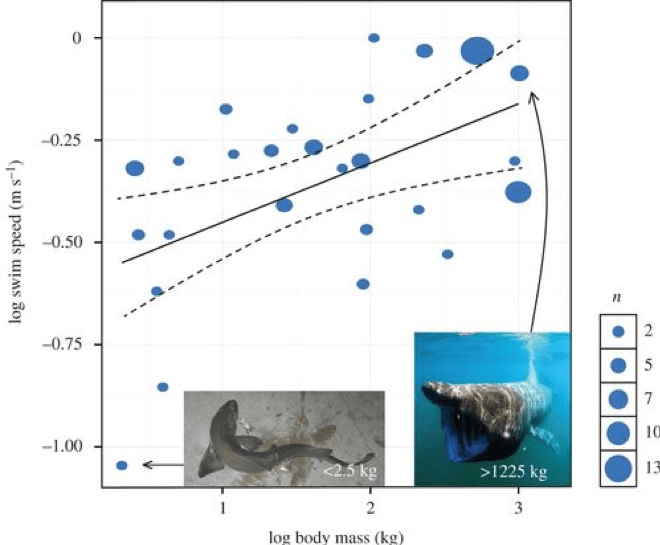
Scientists have also made attempts at calculating Megalodon’s swimming speed, which may help give us an indication of how it hunted. Sharks can have a burst speed, which is a quick jolt of fast swimming, typically used for grabbing prey. But their usual speed for their casual swims are what we call sustained swimming speeds. This varies between species, but one study led by David Jacoby [11] combined data on 26 species from 64 prior studies to develop a model that would predict a scaling relationship between sustained swim speed and the shark’s metabolism and body mass. Their model incorporating metabolism revealed a scaling exponent of 0.173, summarised as the following equation:
Swim Speed (ms ¯¹) ∝ Mass (kg)^0.173
When they corrected the data for phylogeny, however, they found that minimum swim speeds scaled with body mass at the slightly lower exponent of 0.15. The above equation additionally assumes a constant of 1 to match the proportions but the data revealed that this constant was actually 0.266. They therefore constructed the power equation:
Swim Speed (ms ¯¹) = 0.266 Mass (kg) ^0.15
From this, they are able to calculate the sustained swimming speed of Megalodon depending on its mass. For example, the ~48,000 kg Megalodon calculated as the mass of a 15.9 m shark in Gottfried’s work [12] would reveal a shark swimming continuously at 1.34 m/s – roughly 4.8 km/hr [11,13]. Applying this to a 52,000 kg Megalodon reveals a sustained swim speed of 1.36 m/s or 4.9 km/hr whereas if we went really extreme and simulated a 100,000 kg shark, this model would result in a swim speed of 1.50 m/s or 5.4 km/hr. This model has proven very effective not just in calculating Megalodon swim speed, but also in providing a model for multiple shark species (Fig. 4) [11].
Maybe. White shark burst speed is powered by a physiological adaptation called mesothermy [14]. This is something Megalodon has recently been suggested to have had as well (to be discussed in detail in Q5). Mesothermic fishes generally have faster sustained and burst swimming speeds than ectothermic fishes [16], and this has been used to suggest that Megalodon may have had a burst speed of up to 37.15 km/hr [17].
That incredibly quick speed for such a large shark may well be enough to be able to breach. Beyond this, the basking shark (Cetorhinus maximus), today’s second largest shark, has also been observed breaching [18]. Perhaps the giant Megalodon was indeed capable of leaping out of the water as it grabbed its latest meal. We just don’t know yet, but my personal and (somewhat) professional opinion is that it may well have been possible.
With such a broad menu to choose from, Megalodon was an apex predator, with no known predators of its own. This means two important things. Firstly, it likely had stiff competition for its prey, something that may have ultimately contributed to its extinction. Secondly, as an apex predator, Megalodon would’ve been of utmost importance to its ecosystem as it would’ve kept its prey populations in check and thus preserved a healthy ecosystem.
This is why sharks of today are so important to marine environments. The loss of an apex predatory shark produces irreversible cascading effects that change the community it leaves behind [19]. As such, our marine ecosystems are the way they are partially because Megalodon is no longer a part of it. And the enormous whales of modern oceans can rest easy as a result.
References
- Estrada JA, Rice AN, Natanson LJ & Skomal GB 2006. Use of isotopic analysis of vertebrae in reconstructing ontogenetic feeding ecology in white sharks. Ecology 87, 829-834.
- Pimiento C, Ehret DJ, MacFadden BJ & Hubbell G 2010. Ancient nursery area for the extinct giant shark Megalodon from the Miocene of Panama. PLoS One 5, e10552.
- Pimiento C, González-Barba G, Ehret DJ, Hendy AJ, MacFadden BJ & Jaramillo C 2013. Sharks and rays (Chondrichthyes, Elasmobranchii) from the late Miocene Gatun formation of Panama. J. Paleontol. 87, 755-774.
- Purdy RW 1996. Paleoecology of fossil white sharks. In: Great White Sharks: the biology of Carcharodon carcharias (eds. Klimley AP & Ainley DG), 67-78, Academic Press, San Diego.
- Aguilera OA, García L & Cozzuol MA 2008. Giant-toothed white sharks and cetacean trophic interaction from the Pliocene Caribbean Paraguaná Formation. Paläontol. Z. 82, 204-208.
- Fallows C, Gallagher A. & Hammerschlag N 2013. White sharks (Carcharodon carcharias) scavenging on whales and its potential role in further shaping the ecology of an apex predator. PLoS One, 8, e60797.
- Dudley SFJ, Anderson-Reade MD, Thompson GS & McMullen PB 2000. Concurrent scavenging of a whale carcass by great white sharks, Carcharodon carcharias, and tiger sharks, Galeocerdo cuvier. Fish. Bull. 98, 646-649.
- Kallal RJ, Godfrey SJ & Ortner DJ 2010. Bone reactions on a Pliocene cetacean rib indicate short-term survival of predation event. Int. J. Osteoarchaeol. 22, 253–260.
- Collareta A, Lambert O, Landini W, Di Celma C, Malinverno E, Varas-Malca R, Urbina M & Bianucci G 2017. Did the giant extinct shark Carcharocles megalodon target small prey? Bite marks on marine mammal remains from the late Miocene of Peru. Palaeogeogr. Palaeoclimatol. Palaeoecol. 469, 84-91.
- Wroe S, Huber DR, Lowry M, McHenry C, Moreno K, Clausen P, Ferrara TL, Cunningham E, Dean MN & Summers AP 2008. Three‐dimensional computer analysis of white shark jaw mechanics: how hard can a great white bite? J. Zool. 276, 336-342.
- Jacoby DM, Siriwat P, Freeman R & Carbone C 2015. Is the scaling of swim speed in sharks driven by metabolism? Biol. Lett. 11, 20150781.
- Gottfried MD, Compagno LJV & Bowman SC 1996. Size and skeletal anatomy of the giant “megatooth” shark Carcharodon megalodon. In: Great White Sharks: the biology of Carcharodon carcharias (eds. Klimley AP & Ainley DG), 55–66, Academic Press, San Diego.
- Jacoby DM, Siriwat P, Freeman R & Carbone C 2016. Correction to ‘Is the scaling of swim speed in sharks driven by metabolism?’ Biol. Lett. 12, 20160775.
- Watanabe YY, Payne NL, Semmens JM, Fox A & Huveneers C 2019. Swimming strategies and energetics of endothermic white sharks during foraging. J. Exp. Biol. 222, jeb185603.
- Cooper JA, Pimiento C, Ferrón HG & Benton MJ (in press). Body dimensions of the extinct giant shark Otodus megalodon: A 2D reconstruction. Sci. Rep.
- Watanabe YY, Goldman KJ, Caselle JE, Chapman DD & Papastamatiou YP 2015. Comparative analyses of animal-tracking data reveal ecological significance of endothermy in fishes. Proc. Nat. Acad. Sci. 112, 6104-6109.
- Ferrón HG 2017. Regional endothermy as a trigger for gigantism in some extinct macropredatory sharks. PLoS One, 12, e0185185.
- Johnston EM, Halsey LG, Payne NL, Kock AA, Iosilevskii G, Whelan B & Houghton JD 2018. Latent power of basking sharks revealed by exceptional breaching events. Biol. Lett. 14, 20180537.
- Myers RA, Baum JK, Shepherd TD, Powers SP & Peterson CH 2007. Cascading effects of the loss of apex predatory sharks from a coastal ocean. Science, 315, 1846-1850.
Edited by Rhys Charles