
Guest Author: Jack Cooper, MSc
UoB Graduate / PhD Student, University of Swansea
This blog will be discussing some particularly clever recent work on Megalodon. And with it comes a new main player in this ball pit of research. Enter Humberto Ferrón, who I was lucky enough to get to work with on my MSc thesis [1]. He made quite a splash in 2017 when he published two papers of immediate relevance to Megalodon that provided an extremely plausible explanation as to how such a massive shark had managed to evolve in the first place.
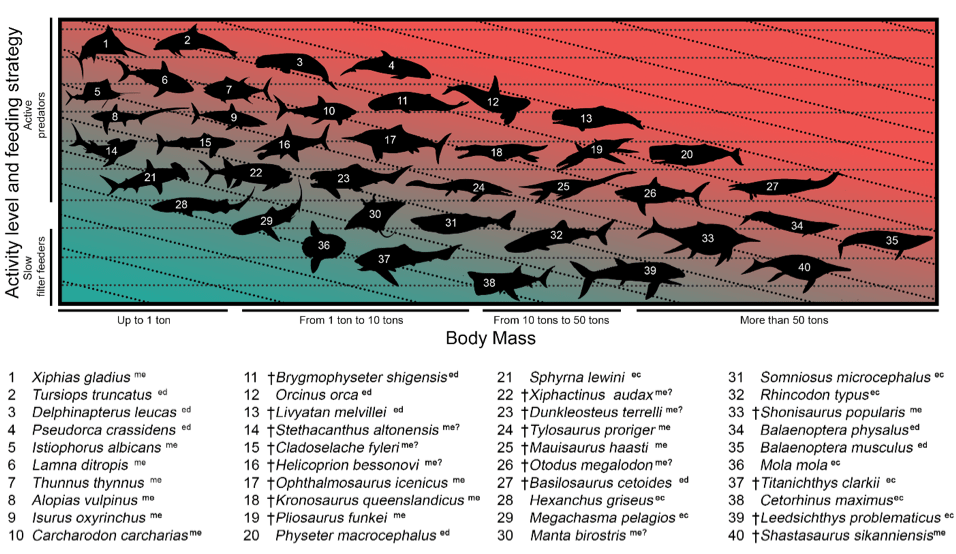
Previous work on body size in marine vertebrates had revealed that energetics was a limiting factor in growing to large sizes. A more active lifestyle chasing prey requires much more metabolic demand that can’t afford to be wasted on getting big. Most of the largest species are thus slow filter feeders, cruising around swallowing plankton.
However, Ferrón and colleagues Carlos Martinez-Pérez and Héctor Botella noticed that there was a clear size difference between the largest marine vertebrates of two differing thermoregulatory strategies regardless of the feeding ecology involved [2]. Among the largest filter feeders, the blue whale (Balaenoptera musculus) is endothermic (able to internally maintain body temperature via metabolic processes) and more than 10 m larger than the whale shark (Rhincodon typus), which is ectothermic and therefore relies on its environment to maintain its body temperature. Meanwhile, in active macropredators with higher metabolic levels, the endothermic sperm whale (Physeter macrocephalus) is quadruple the size of the ectothermic Greenland shark (Somniosus microcephalus).
Activity and feeding strategies in living organisms are constrained by mass-specific metabolic rates. This typically decreases with increasing body size, meaning that it’s better for a big animal to conserve energy by becoming a slow filter feeder. However, this limit appeared to be reached at different masses depending on the thermoregulatory strategy. As such, endotherms whose body temperatures remain fairly stable regardless of environmental temperature could get larger before hitting their metabolic limits. Ferrón and his colleagues published this novel hypothesis, providing a widespread explanation for the presence of some truly humongous active marine predators in geologic history, including Dunkleosteus, mosasaurs and, of course, Megalodon (Fig. 1) [2].
Having any kind of independent thermoregulation is unusual in sharks, most of which are ectothermic [5]. As such, most early researchers had simply assumed that Megalodon and its megatoothed relatives were also ectothermic. However, Ferrón noted that this had been challenged more recently. In his PhD thesis [6], Dana Ehret had questioned this as he had found no apparent correlation between Megalodon’s growth rate and known Miocene and Pliocene sea temperatures. He therefore suggested that perhaps Megalodon had some kind of endothermic adaptation. Catalina Pimiento had also put forward this possibility as her paper on Megalodon’s global occurrences had found that it occupied a wide range of water temperatures [7]. Ferrón therefore proposed that mesothermy was a trait in the shared common ancestor of Megalodon and lamnid sharks like the great white [3]. Dana Ehret’s PhD thesis suggested that this was rather parsimonious (the simplest explanation for its presence in both groups). This ancestor likely dated back to the Cretaceous, so Ferrón set out to test this physiological hypothesis directly.
He did this through three separate methods. Firstly, he analysed oxygen isotopes of the fossils’ phosphate deposits. This had previously been done for Mesozoic marine reptiles and confirmed that some of them had used endothermy [8]. By comparing the results of his fossils to known ectotherms from the same formations, Ferrón could uncover a difference in body temperature despite living in the same environment. Indeed he did find this for Megalodon and its relatives [3].
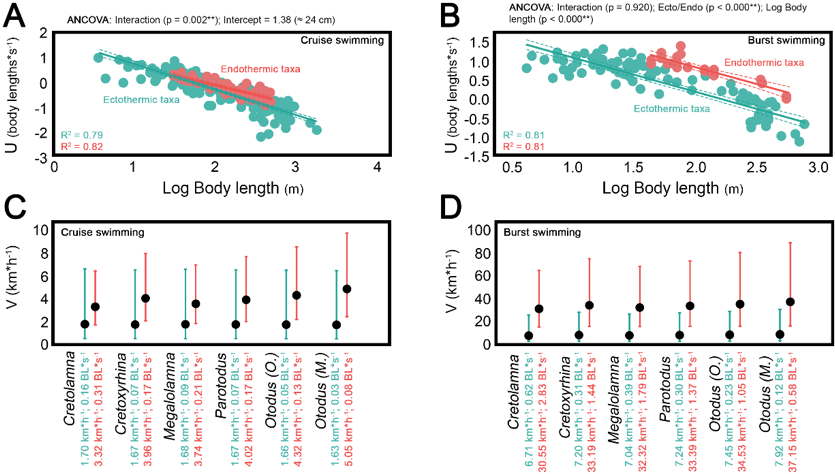
Secondly, he assessed if burst swimming speeds would be more successful as an endotherm or an ectotherm for these giant sharks. He did this by compiling swimming and thermoregulatory data of extant shark species to compare cruising and burst speeds in ectotherms and endotherms (Fig. 2). His findings agreed with previous work [9] that cruising speeds of endothermic sharks were greater than those of ectotherms, and that mesothermy could produce a higher burst speed – meaning that the mesothermic Megalodon could strike quickly when it came across prey, something that the great white notably uses to catch seals [3,9,10].
Specifically, he found that Megalodon crusing speed was around 5.05 km/hr – similar to previous suggestions of 1.34 m/s [11,12] – and had a potential burst speed of up to 37.15 km/hr [3]. Such speeds were only possible as a result of mesothermy.
Finally, he found that Cretoxyrhina mantelli (a possible common ancestor between Megalodon and lamnid sharks) had the same basic tail shape as lamnids [9,13] – something that he and I (alongside Catalina Pimiento and Bristol’s own Mike Benton) would together suggest was also used by Megalodon in my MSc thesis [1]. This is perfect for fast, efficient swimming that would only be possible for such a large shark if endothermic in some way. Therefore, Ferrón concluded that the megatoothed clade, including Megalodon, was likely mesothermic, and that this directly contributed to its ginormous size [3].
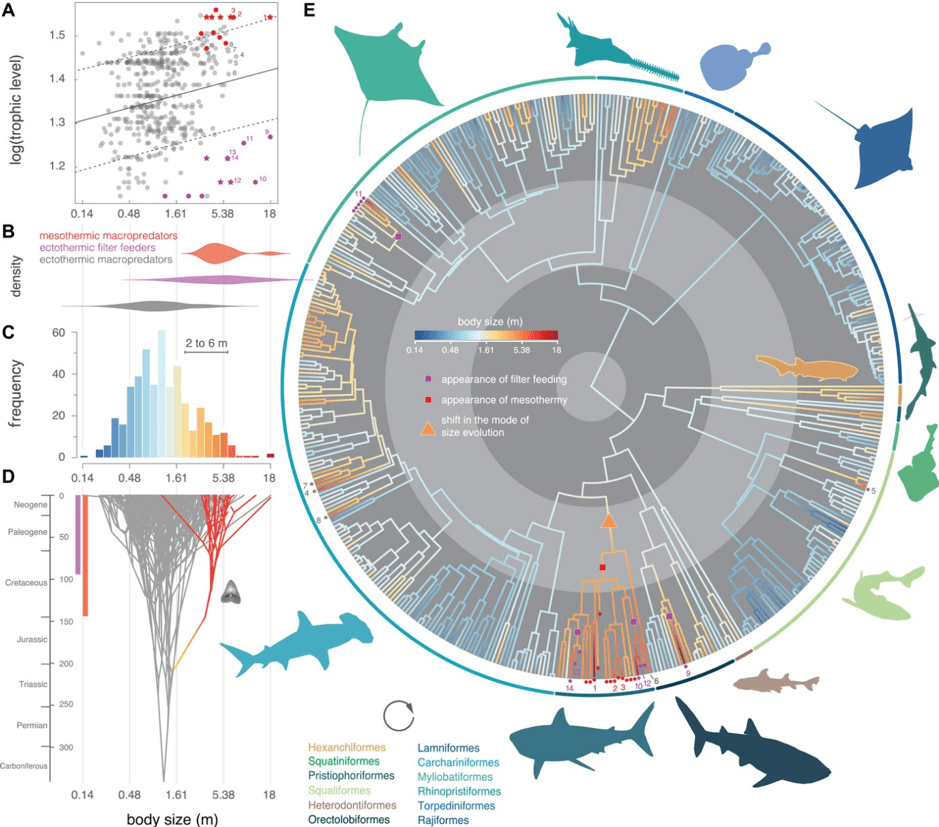
By running a phylogenetic analysis, they uncovered that gigantism had evolved independently several times in sharks (Fig. 3). However, while ectothermic macropredators only reached a maximum size of 7.5 m TL, filter feeders and mesothermic macropredators could both reach 18 m TL (whale shark and Megalodon respectively) (Fig. 3).
This strongly suggested that one of these strategies was needed for a shark to achieve gigantism. Digging deeper into the origins of these pathways, Pimiento and her team found that filter feeding had evolved independently four times in sharks while mesothermy could be traced back to the Cretaceous, around the time C. mantelli was just evolving. They therefore theorised that gigantism was not an ancestral trait to any shark, instead a result of either the filter feeding or mesothermy adaptations [14]. As such, with Megalodon clearly not being a filter feeder, it seems that this physiological adaptation brought to wider attention by Humberto Ferrón was indeed the trigger for Megalodon to become a giant.
References
- Cooper JA, Pimiento C, Ferrón HG & Benton MJ (in press). Body dimensions of the extinct giant shark Otodus megalodon: A 2D reconstruction. Sci. Rep.
- Ferrón HG, Martínez-Pérez C & Botella H 2017. The evolution of gigantism in active marine predators. Hist. Biol. 30, 712-716.
- Ferrón HG 2017. Regional endothermy as a trigger for gigantism in some extinct macropredatory sharks. PLoS One, 12, e0185185.
- Compagno LJV 2001. FAO Species catalogue. Sharks of the world. An annotated and illustrated catalogue of shark species known to date. FAO Species Catalogue Fishery Purposes 2.
- Dickson KA & Graham JB 2004. Evolution and consequences of endothermy in fishes. Physiol. Biochem. Zool. 77, 998-1018.
- Ehret DJ 2010. Paleobiology and taxonomy of extinct lamnid and otodontid sharks (Chondrichthyes, Elasmobranchii, Lamniformes). PhD thesis, University of Florida.
- Pimiento C, MacFadden BJ, Clements CF, Varela S, Jaramillo C, Velez‐Juarbe J & Silliman BR 2016. Geographical distribution patterns of Carcharocles megalodon over time reveal clues about extinction mechanisms. J. Biogeogr. 43, 1645-1655.
- Bernard A, Lécuyer C, Vincent P, Amiot R, Bardet N, Buffetaut E, Cuny G, Fourel F, Martineau F, Mazin JM & Prieur A 2010. Regulation of body temperature by some Mesozoic marine reptiles. Science, 328, 1379-1382.
- Watanabe YY, Goldman KJ, Caselle JE, Chapman DD & Papastamatiou YP 2015. Comparative analyses of animal-tracking data reveal ecological significance of endothermy in fishes. Proc. Nat. Acad. Sci. 112, 6104-6109.
- Watanabe YY, Payne NL, Semmens JM, Fox A & Huveneers C 2019. Swimming strategies and energetics of endothermic white sharks during foraging. J. Exp. Biol. 222, jeb185603.
- Jacoby DM, Siriwat P, Freeman R & Carbone C 2015. Is the scaling of swim speed in sharks driven by metabolism? Biol. Lett. 11, 20150781.
- Jacoby DM, Siriwat P, Freeman R & Carbone C 2016. Correction to ‘Is the scaling of swim speed in sharks driven by metabolism?’ Biol. Lett. 12, 20160775.
- Kim SH, Shimada K & Rigsby CK 2013. Anatomy and evolution of heterocercal tail in lamniform sharks. Anat. Rec. 296, 433-442.
- Pimiento C, Cantalapiedra JL, Shimada K, Field DJ & Smaers JB 2019. Evolutionary pathways toward gigantism in sharks and rays. Evolution, 73, 588-599.
Edited by Rhys Charles